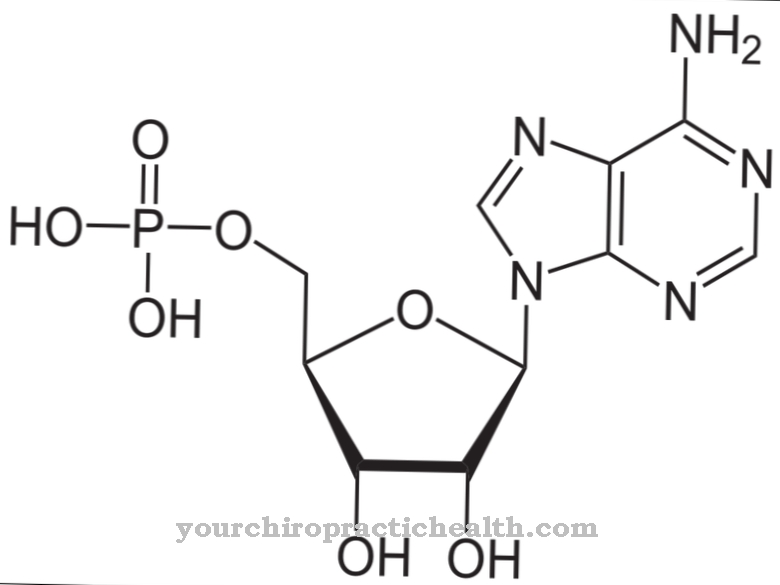
nukleobaser er byggesteinene som de lange kjedene av DNA og RNA-molekyler er sammensatt av i deres fosforylerte nukleotidform.
I DNAet, som danner doble tråder som ligner tau, danner de 4 forekommende nukleiske baser faste parringer med den respektive komplementære basen via hydrogenbroer. Nukleobasene består av enten et bicyklisk purin eller et monocyklisk pyrimidinskjelett.
Hva er nukleobaser?
De 4 nukleobaser adenin, guanin, cytosin og tymin, som byggesteiner i de lange DNA-dobbelhelixmolekylkjedene, danner de konstante sammenkoblinger adenintymin (A-T) og guanin-cytosin (G-C).
De to basene adenin og guanin består hver av en modifisert bicyklisk seks- og femdelersring av den grunnleggende purinstrukturen og blir derfor også referert til som purinbaser. Grunnstrukturen til de to andre nukleiske basene, cytosin og timin, består av en heterocyklisk aromatisk sekstremsring, som tilsvarer et modifisert pyrimidinskjelett, og det er derfor de også omtales som pyrimidinbaser. Siden RNA stort sett er til stede som enkeltstrenger, er det i utgangspunktet ingen basepar. Dette skjer bare under replikering via mRNA (messenger RNA).
Kopien av RNA-strengen består av de komplementære nukleobaser som er analoge med den andre DNA-strengen. Den eneste forskjellen er at uracil er erstattet med timin i RNA. DNA- og RNA-kjedemolekylene er ikke dannet i sin rene form av nukleobasene, men snarere, i tilfelle av DNA, kombineres de med 5-sukker deoxyribose for å danne det tilsvarende nukleosid. For RNA består sukkergruppen av ribose. I tillegg fosforyleres nukleosidene med en fosfatrest for å danne såkalte nukleotider.
Purinbasene hypoksantin og xantin, som også forekommer i DNA og RNA, tilsvarer modifisert timin. Hypoksantin dannes fra adenin ved å erstatte aminogruppen (-NH3) med en hydroksylgruppe (-OH), og xantin dannes fra guanin. Begge nukleobaser bidrar ikke til overføring av genetisk informasjon.
Funksjon, effekt og oppgaver
En av de viktigste funksjonene til nukleobasene, hvorfra de doble DNA-strengene er bygget, er å vise tilstedeværelse på den tiltenkte posisjonen.
Sekvensen til nukleobasene tilsvarer den genetiske koden og definerer typen og sekvensen av aminosyrer som proteiner er sammensatt av. Dette betyr at den viktigste funksjonen til nukleobasene som en del av DNAet består av en passiv, statisk rolle, dvs. at de ikke aktivt griper inn i metabolismen og deres biokjemiske struktur blir ikke endret av messenger RNA (mRNA) under leseprosessen. Det forklarer delvis DNA-levetiden.
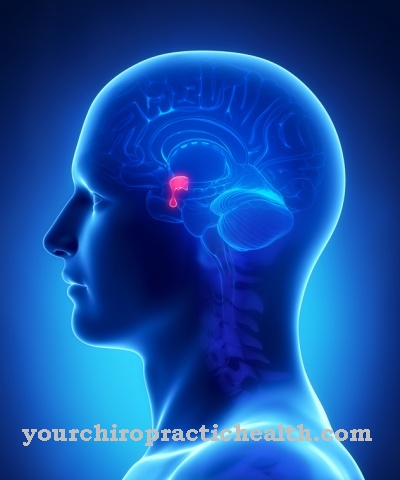
Halveringstiden for mitokondrielt DNA (mtDNA), der halvparten av de opprinnelig eksisterende bindingene mellom nukleobasene går i oppløsning, er sterkt avhengig av miljøforholdene og varierer mellom 520 år under gjennomsnittlige forhold med positive temperaturer og opptil 150 000 år under permafrostforhold .
Som en del av RNA spiller nukleobasene en noe mer aktiv rolle. Når celler deler seg, blir DNA-dobbeltrådene i prinsippet brutt opp og separert fra hverandre for å kunne danne en komplementær streng, mRNA, som så å si danner arbeidskopien til det genetiske materialet og fungerer som grunnlag for seleksjonen og sekvensen av aminosyrene som tiltenkte proteiner er samlet. En annen nukleisk base, dihydrouracil, forekommer bare i den såkalte transport-RNA (tRNA), som brukes til å transportere aminosyrer under proteinsyntese.
Noen nukleobaser har en helt annen funksjon som en del av enzymer som aktivt katalytisk muliggjør og kontrollerer visse biokjemiske prosesser. Adenin oppfyller sin mest kjente oppgave som nukleotid i energibalansen til celler. Her spiller adenin en viktig rolle som elektronisk donor som adenosindifosfat (ADP) og adenosintrifosfat (ATP), så vel som en komponent av nikotinamid adenindinukleotid (NAD).
Utdanning, forekomst, egenskaper og optimale verdier
I den ikke-fosforylerte formen består nukleobaser utelukkende av karbon, hydrogen og oksygen, stoffer som er allestedsnærværende og fritt tilgjengelige. Kroppen er derfor i stand til å syntetisere nukleobaser selv, men prosessen er kompleks og energikrevende.
Derfor foretrekkes utvinning av nukleinsyrer ved resirkulering, f.eks. B. ved å bryte ned proteiner som inneholder visse forbindelser som kan isoleres og omdannes til nukleinsyrer med lite energiforbruk eller til og med med energigevinst. Nukleinsyrer forekommer vanligvis ikke i ren form i kroppen, men mest som nukleosider eller deoksynukleosider med en festet ribose- eller deoksyribosemolekyl. Som en komponent av DNA og RNA og som en komponent i visse enzymer, blir nukleinsyrene eller deres nukleosider også reversibelt fosforylert med en til tre fosfatgrupper (PO4-).
Det er ingen referanseverdi for en optimal tilførsel av nukleobaser. En mangel eller et overskudd av nukleobaser kan bare bestemmes indirekte via visse forstyrrelser i metabolismen.
Sykdommer og lidelser
Den typen farer, forstyrrelser og risikoer som bringes i forbindelse med nukleobasene er feil i antall og sekvens på DNA- eller RNA-strengene, noe som fører til en endring i kodingen for proteinsyntese.
Hvis kroppen ikke kan avhjelpe feilen via sine reparasjonsmekanismer, kommer det til syntese av biologisk inaktive eller anvendbare proteiner, som igjen kan føre til milde til alvorlige metabolske forstyrrelser. Det kan f.eks. B. genetiske mutasjoner er til stede som kan utløse symptomatiske sykdommer fra begynnelsen via metabolske forstyrrelser, som kan være uhelbredelige. Men selv i et sunt genom kan det oppstå kopieringsfeil under replikasjonen av DNA- og RNA-kjedene, som påvirker stoffskiftet.
En kjent metabolsk lidelse i purinbalansen er z. B. tilbake til en genetisk defekt på x-kromosomet. På grunn av den genetiske defekten, kan ikke purinbasen hypoksantin og guanin gjenvinnes, noe som til slutt fremmer dannelsen av urinstein og gikt i leddene.





.jpg)



.jpg)