De Membranpermeabilitet karakteriserer permeabiliteten til molekyler gjennom cellemembranen. Alle celler skilles fra det intercellulære rommet med biomembraner og inneholder samtidig celleorganeller som igjen er omgitt av membraner. Gjennomtrengeligheten av membranene er nødvendig for en jevn drift av de biokjemiske reaksjonene.
Hva er membranpermeabiliteten?

Membranpermeabiliteten er definert som permeabiliteten til biomembranen for væsker og oppløste stoffer. Cellemembraner er imidlertid ikke gjennomtrengelige for alle stoffer. Derfor blir de også referert til som semipermeable membraner (halvgjennomtrengelige membraner).
Biomembraner består av to fosfolipidlag som er gjennomtrengelige for gasser som oksygen eller karbondioksid samt lipidløselige, ikke-polare stoffer. Disse stoffene kan passere gjennom membranene via normal diffusjon. Polare og hydrofile molekyler er ikke tillatt. De kan bare transporteres gjennom membranen via passive eller aktive transportprosesser.
Membraner beskytter det indre-celleformede rommet og rommet i celleorganellene. De sikrer opprettholdelse av spesielle kjemiske og fysiske forhold for viktige biokjemiske reaksjoner uten ytre forstyrrelser.
Membranenes permeabilitet sikrer selektiv transport av viktige stoffer fra det ekstracellulære rommet inn i cellen og fjerning av metabolske produkter fra cellen. Det samme gjelder de individuelle celleorganellene.
Funksjon & oppgave
Membraner er helt nødvendige for den uforstyrrede prosessen med viktige biokjemiske reaksjoner i cellene og celleorganellene. Membranpermeabiliteten er like viktig for å kunne forsyne cellene med viktige næringsstoffer som proteiner, karbohydrater eller fett. Mineraler, vitaminer og andre aktive ingredienser må også kunne passere gjennom membranen. Samtidig lages metabolske produkter som må kastes fra cellen.
Imidlertid er membranene bare gjennomtrengelige for lipofile molekyler og små gassmolekyler som oksygen eller karbondioksyd. Polare, hydrofile eller store molekyler kan bare transporteres gjennom membranen ved hjelp av transportprosesser. Det er passive og aktive alternativer for membrantransport for dette.
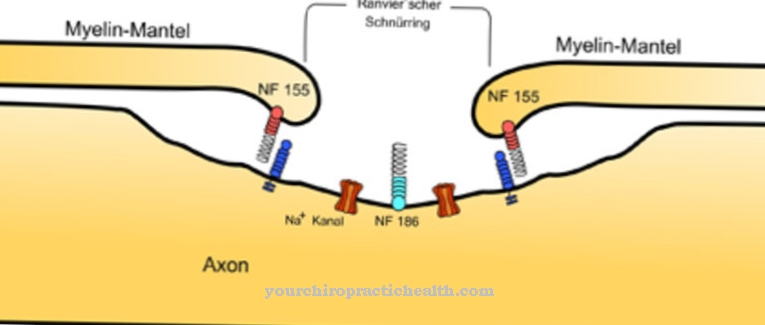
Passiv transport fungerer uten å tilføre energi i retning av en potensiell eller konsentrasjonsgradient. Mindre lipofile molekyler eller gassmolekyler er utsatt for normal diffusjon. Normal diffusjon er ikke lenger mulig med større molekyler. Visse transportproteiner eller kanalproteiner kan lette transporten hit. Transportproteinene spenner over membranen som en tunnel. Mindre polare molekyler kan føres gjennom denne tunnelen via virkningen av polare aminosyrer. Dette muliggjør også transport av små ladede ioner gjennom tunnelen.
Et annet passivt transportalternativ er resultatet av virkningen av bærerproteiner som spesialiserer seg i visse molekyler. Når molekylet la til kai, endrer de sin konformasjon og transporterer det gjennom membranen.
Ved aktiv membrantransport er tilførsel av energi nødvendig. Det tilsvarende molekylet transporteres mot en konsentrasjonsgradient eller en elektrisk gradient. Energiforsyningsprosesser er resultatet av hydrolyse av ATP, oppbygging av en ladningsgradient i form av et elektrisk felt eller økningen i entropi ved å bygge opp en konsentrasjonsgradient.
Endocytose eller eksocytose er tilgjengelig for stoffer som ikke kan trenge gjennom membranen i det hele tatt. Ved endocytose tar invaginasjonen av biomembranen en dråpe væske og transporterer den inn i cellen. Dette skaper et såkalt endosom, som transporterer viktige stoffer til cytoplasma. Under eksocytose blir avfallsprodukter i cytoplasma transportert utover med membranbelagte transportvesikler.
Sykdommer og plager
Forstyrrelser i membranpermeabilitet kan føre til forskjellige sykdomstilstander. Endringene påvirker permeabiliteten til de forskjellige ionene. Membranpermeabilitetsforstyrrelser er ofte et resultat av hjerte- og karsykdommer. Dette kan påvirke kroppens elektrolyttbalanse.
Imidlertid forårsaker mange arvelige årsaker også membranpermeabilitetsforstyrrelser. Ulike proteiner er involvert i strukturen av membranen og er ansvarlige for riktig funksjon av lipid-dobbeltlaget. Genetiske forandringer i visse proteiner er blant annet ansvarlige for endringer i membranpermeabilitet.
Et eksempel er sykdommen Myotonia congenita Thomsen. Denne sykdommen er en genetisk forstyrrelse av muskelfunksjon. Et gen som koder for kloridkanalene til muskelfibermembraner er mutert. Kloridionens permeabilitet reduseres. Dette fører til en enklere depolarisering av muskelfibre enn hos friske mennesker. Tendensen til muskelsammentrekning økes, noe som kjennes som stivhet. For eksempel kan en lukket knyttneve bare åpnes med en viss forsinkelse. Øynene kan bare åpnes etter 30 sekunder etter lukking, som er kjent som lokk-lag.
Det er også autoimmune sykdommer som spesifikt retter seg mot biomembraner. I denne sammenhengen er det såkalte antifosfolipid syndrom (APS) kjent. I denne sykdommen er kroppens immunsystem rettet mot proteiner som er bundet til fosfolipidene i membranen. Resultatet er en økt koagulerbarhet i blodet. Sannsynligheten for hjerteinfarkt, hjerneslag og lungeembolismer er økt.
Forstyrrelser i membranpermeabilitet kan også finnes i den såkalte mitokondriesykdommen. I mitokondriene oppnås energi fra forbrenning av karbohydrater, fett og proteiner. Mitokondriene er celleorganeller som også er omgitt av en membran. Et stort antall frie radikaler produseres i disse energikraftverkene. Hvis disse ikke fanges opp, vil membranene bli skadet. Dette begrenser mitokondriens funksjon sterkt. Årsakene til den reduserte effektiviteten til de radikale oppfangerne er imidlertid forskjellige.


.jpg)

.jpg)

.jpg)




.jpg)



.jpg)