Av passiv massetransport er diffusjon av underlag gjennom en biomembran. Denne diffusjonen skjer langs konsentrasjonsgradienten og krever ingen energi. Diffusjonsprosessen kan for eksempel forstyrres i tarmen til HIV-pasienter.
Hva er passiv massetransport?
![]()
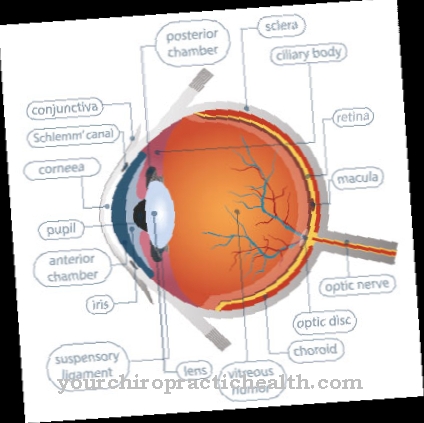
Celler eller celleformasjoner skilles fra hverandre i kroppen med en biomembran. Takket være sine spesialiserte strukturer muliggjør dette fleksible skillesjiktet transport av spesifikke molekyler og informasjon inn og ut av celleinteriøret.
Det er to grunnleggende måter å transportere stoffer inn og ut av membranen. Membraner har en selektiv permeabilitet. De lar noen stoffer diffundere mens de fungerer som en barriere for andre.
Den aktive transporten av stoffer betyr at membraner kan åpne på en målrettet måte for molekyler som de faktisk ikke er gjennomtrengelige på grunn av deres ladning, konsentrasjon eller størrelse. Aktiv transport skjer alltid med bruk av energi. Det skal skilles mellom denne og typen massetransport av passiv transport. Det kreves ingen energi i denne formen for bevegelse av materie gjennom en cellemembran. Passiv transport skal likestilles med diffusjonsprosesser som foregår langs konsentrasjonsgradienten og gir en konsentrasjonsbalanse mellom membranens to sider.
Funksjon & oppgave
I en celle eller et cellerom er det et visst kjemisk og ladende miljø som er nødvendig for at cellen skal fungere. Dette miljøet opprettholdes bare av egenskapene til biomembranen og den selektive permeabiliteten. Den passive og aktive substanstransporten forsyner cellen eller cellerommet i nøyaktig riktig mengde med nøyaktig de stoffene som er nødvendige for et gunstig miljø.
Det er to forskjellige typer passiv transport. Den enkle diffusjonen påvirker lipidløselige molekyler og forekommer i en ekstremt langsom hastighet. De diffunderer fritt gjennom cellemembranen. Denne formen for passiv transport er den som har minst krefter. Den andre typen passiv diffusjon er tilrettelagt for diffusjon, som igjen kan deles i to underformer. En av disse underformene er bærermediert forenklet diffusjon. Med denne formen for passiv masseoverføring henter membranen underlaget ved hjelp av en såkalt bærer. Bæreren er et protein som identifiserer stoffet som underlaget binder seg til. Siden enkel diffusjon skjer sakte, hjelper bæreren med å transportere stoffet gjennom biomembranen. Antallet av bærermolekyler er begrenset.
Av denne grunn er transporten gjennom et bærermolekyl utsatt for metningskinetikk. Passiv transport av stoffer med bærermolekyler kan også bli utsatt for konkurrerende hemming. Når et bærermolekyl kobles til underlaget, endrer det sin konformasjon og omorganiseres deretter. Som et resultat blir substratmolekylet transportert gjennom biomembranen og bare frigjort igjen på motsatt side.
Noen bærere kan bare bære ett molekyl av gangen og dermed ha en uniport. Andre bærere har bindingssteder for to forskjellige molekylære underlag og endrer kun konformasjonen når begge bindingsseter er okkupert. De to molekylene er enten symport i samme retning eller antiport i motsatte retninger. Det er ingen avhengighet av den elektriske gradienten.
Den andre typen forenklet diffusjon er gjennom porer og kanaler. Denne transportformen påvirker spesielt aminosyrer. Under ionetransport blir for eksempel aminosyrens underlag absorbert i cellemembranen gjennom porene. Kanalene dannes av proteiner. Det er spesielle bindingssteder på disse proteinholdige kanaler. Den tilrettelagte diffusjonen gjennom porer og kanaler er en selektiv materialtransport som kan påvirkes elektrisk og kjemisk.
Nesten alle kanaler åpnes bare som svar på visse signaler. For eksempel reagerer en ligandstyrt kanal bare på et messenger-stoff som et hormon. Noen kanaler er spenningskontrollerte og åpne for diffusjon med endring i membranpotensial. Etter konsentrasjonsutjevningen lukkes kanalene igjen.
Sykdommer og plager
Hvis membranpermeabiliteten og dermed også den passive massetransporten blir forstyrret, er permeabiliteten til forskjellige ioner ikke lenger ideell regulert. Slike membranpermeabilitetsforstyrrelser utvikler seg ofte fra hjerte- og karsykdommer og svekker noen ganger elektrolyttbalansen.
Noen ganger er membranpermeabilitetsforstyrrelser også arvelige. Ulike proteiner bygger opp biomembranen og gir den et selektivt permeabelt dobbelt lipidlag. Når de involverte proteinene endres, endres også membrangjennomtrengeligheten. Dette fenomenet forekommer for eksempel i Myotonia congenita Thomsen. Denne genetiske forstyrrelsen i muskelfunksjon får et gen til å mutere som er ansvarlig for koding av de individuelle kloridkanalene i muskelfibermembranene. På grunn av mutasjonen reduseres permeabiliteten for kloridioner og forårsaker dermed muskelstivhet.
Autoimmune sykdommer kan også rettes mot biomembranen, for eksempel antifosfolipidsyndromet. Immunsystemet angriper fosfolipidbundne proteiner i membranen som en del av sykdommen. Den økte tendensen til blodpropp øker også risikoen for hjerteinfarkt og hjerneslag.
Mitokondriesykdommer endrer også permeabiliteten til membranene. Mitokondriene er kroppens egne energikraftverk, som frigjør frie radikaler når de genererer energi. Disse stoffene blir oppfanget hos friske mennesker. Denne prosessen mislykkes hos pasienter med mitokondiopati, noe som skader membranene og reduserer mitokondriens evne til å produsere energi i stor grad.
Den passive og aktive transporten av stoffer gjennom tynntarmens membraner påvirkes spesielt av lidelser som HIV-enteropati. Dette fenomenet rammer spesielt HIV-pasienter med kronisk diaré og kan være assosiert med redusert aktivitet av de interstinale enzymer.












.jpg)
.jpg)
.jpg)
.jpg)