Gjennom Saltende ledning av eksitasjon den tilstrekkelig raske konduksjonshastigheten til nervesystemer er sikret for virveldyr. Handlingspotensialer hopper fra den ene uisolerte ringen til den neste på de isolerte aksonene. Ved demyeliniserende sykdommer brytes isolerende myelin ned, noe som forstyrrer ledning av eksitasjon.
Hva er saltdannende ledning av eksitasjon?

Den saltdannende ledningen av eksitasjon er en form for nerveledningsevne. I organismen av virveldyr blir nervefibrene elektrisk isolert fra omgivelsene ved myelinskjede og påtar seg dermed funksjonen til en kappet kabel. Excitasjonen av en nervefiber skjer ved avbrudd i dette isolerende laget, som også er kjent som bindinger eller knuter.
Mange nervefibre i virveldyr har tynn form. Tynne aksoner har lavere ledningshastighet enn sterke nervesystemer. Slik at ledningshastigheten til nervene er tilstrekkelig til tross for den lave styrken, er eksitasjonsledningen til virveldyrene bygd opp salting og bruker både biokjemiske og bioelektriske prosesser for å overføre handlingspotensialer.
Handlingspotensialet hopper fra den ene ringen til den andre i denne typen ledning og etterlater de omhylmede delene av aksonene. En høyere ledningshastighet oppnås med dette prinsippet gjennom spenningsavhengige natriumpumper og bioelektriske biokjemiske prosesser.
Funksjon & oppgave
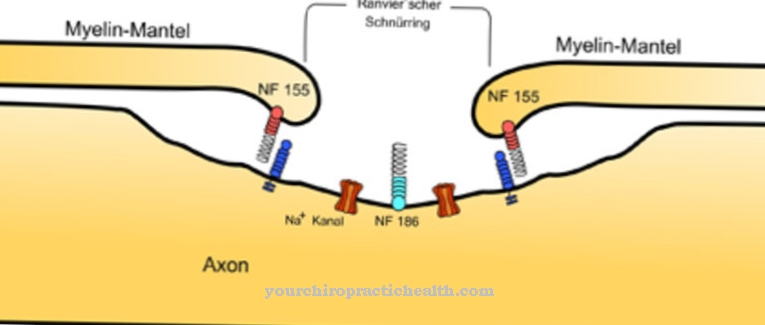
I det perifere nervesystemet danner Schwanns celler myelin som skjeder nervene. Oligodendrocytter påtar seg denne oppgaven i sentralnervesystemet. Aksoner i begge systemer er belagt med myelin, som har en elektrisk isolerende effekt. Isolasjonen av aksonene blir avbrutt i en avstand mellom 0,2 og 1,5 millimeter. Disse avbruddene er også kjent som knuter eller Ranvier-bånd. I kontrast kalles de myelinbelagte seksjonene internoder og sikrer en redusert membran tidskonstant, noe som sikrer en ledningshastighet på 100 meter per sekund. Det er også spenningsavhengige natrium + -kanaler i de glideløse snøringsringene.
Så lenge et akson ikke er begeistret, råder det såkalte hvilepotensialet i sin node og langs sin internode. Mellom det intracellulære rommet og det ekstracellulære rommet til aksonet er det en potensiell forskjell med hvilepotensialet. Når et handlingspotensial blir generert på den første kjeglen til eksitasjonslinjen, som depolariserer dens membran utover dens terskelpotensial, åpnes de spenningsavhengige Na + -kanalene. På grunn av elektrokjemiske egenskaper strømmer Na + -ioner deretter fra det ekstracellulære rommet inn i det intracellulære rommet.
Plasmamembranen depolariserer ved nivået av kjegleringen, og membrankondensatoren lades innen 0,1 ms. I området med snørringen er det et intracellulært overskudd av positive ladningsbærere sammenlignet med omgivelsene på grunn av natriumionene som har flydd inn. Et elektrisk felt skapes. Dette feltet genererer en potensiell forskjell langs aksonet og har innflytelse på ladede deler i nærmeste avstand.
De negativt ladede partiklene på neste ring tiltrekkes av den overskytende positive ladningen i den første ringen. Positivt ladede partikler mellom den første og andre innsnevringsringer beveger seg mot den andre noden. Disse ladningsforskyvningene påvirker membranpotensialet til den andre kjegleringen positivt, selv om ionene ikke har nådd den. På denne måten hopper eksitasjonen fra ring til ring og beholder egenskapen til å depolarisere membranen til de etterfølgende ringer tilstrekkelig.
Sykdommer og plager
Demyeliniserende sykdommer bryter ned myelinskjeder rundt nervefibrene. Disse myelin-skjede er en forutsetning for saltvannsledelse av eksitasjon. Uten myelinhylsen oppstår høye strømtap i internoden. Derfor kreves det større eksitasjoner slik at aksonene kan depolarisere de neste båndringene via et handlingspotensial.
Som regel er handlingspotensialet som overføres etter tapene for lavt til å bli gjenkjent som sådan ved neste node. Som et resultat viderefører ikke snørringen spenningen.
Fenomenet demyelinering er også kjent som demyelinisering og hører til degenerative sykdommer. Aldersrelaterte prosesser så vel som giftige og inflammatoriske prosesser kan avmarkere aksonene og dermed sette den saltvannende overføringen av handlingspotensialer i fare.
Vitaminmangel kan også være relatert til dette fenomenet. Spesielt for lite vitamin B6 og vitamin B12 er assosiert med demarking. En slik vitaminmangel finnes ofte i alkoholisme, for eksempel. En demyelinisering av nervesystemet kan også skje i sammenheng med narkotikamisbruk.
Den mest kjente inflammatoriske årsaken til misfarging av nervene er den autoimmune sykdommen multippel sklerose. Det eget immunsystemet ødelegger nervevevet i sentralnervesystemet som en del av sykdommen. Andre årsaker til demarking kan være diabetes, Lyme sykdom eller genetiske sykdommer. De genetiske sykdommene med demyeliniserende egenskaper inkluderer for eksempel Krabbes sykdom, Pelizaeus-Merzbachers sykdom og Déjérine-Sottas syndrom.
Symptomene som oppstår ved avmyelinisering av nervevevet, avhenger av plasseringen av de avyeliniserende fociene. I sentralnervesystemet kan for eksempel avyelinisering føre til svekkelse av sanseorganene, spesielt til svekkelse i øynene. Lammelse er også tenkelig i tilfelle av demyelinisering i sentralnervesystemet, siden motoriske nervesystemer og kontrollsentrene deres befinner seg der. I det perifere nervesystemet er avyelinisering av nervene sjeldnere forbundet med lammelse. På den annen side kan demyelinisering av perifere aksoner føre til nummenhet eller andre sensoriske lidelser.
Diagnosen avyeliniserende sykdom stilles ved bruk av bildeteknikker som magnetisk resonansavbildning. MR-bildene viser typisk hvite focier av demyeliniserende når kontrastmiddel administreres.



.jpg)

.jpg)

.jpg)




.jpg)



.jpg)