De komplekse cellulære og fysiologiske prosessene i levende vesener krever finstemt regulering på molekylært nivå for å sikre tilpasningsevnen til et dyr eller en plante, for eksempel til habitatet. For dette formålet er det mange molekyler som griper inn i prosesser som cellekommunikasjon, metabolisme eller celledeling. Et av disse molekylene er protein calmodulinsom ved hjelp av kalsium påvirker funksjonen til mange andre biologisk aktive proteiner.
Hva er calmodulin?
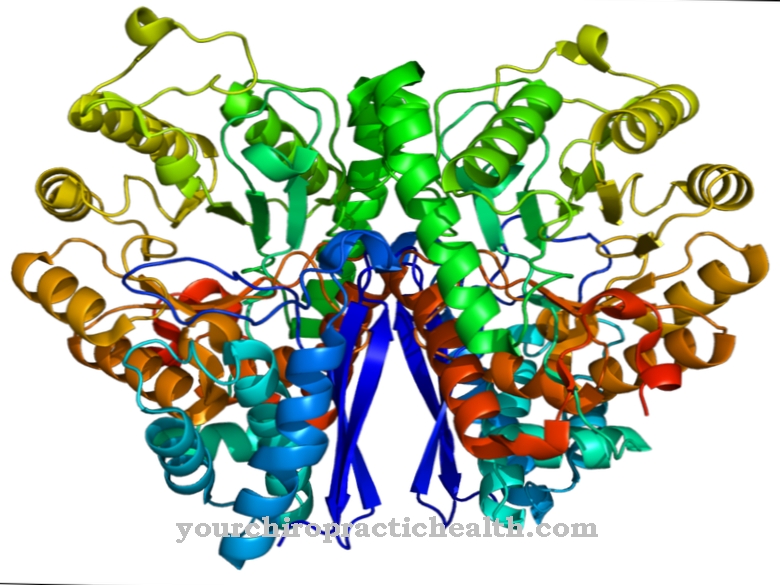
Calmodulin er et intracellulært regulatorisk protein som binder kalsiumioner. På grunn av strukturen tilhører den gruppen av EF-håndproteiner. Formen på calmodulin, som består av 148 aminosyrer og er 6,5 nm lang, ligner en hantel. Molekylmassen til dette proteinmolekylet er omtrent 17 kDa.
På grunn av sin biologiske funksjon i signaloverføring i celler, kan calmodulin også klassifiseres som en andre messenger, dvs. et sekundært messenger-stoff som imidlertid ikke selv er enzymatisk aktivt. I de to sfæriske domenene til proteinet er det to helix-loop-helix-motiver i en avstand på 1,1 nm, til hvilke totalt fire kalsiumioner kan bindes. Denne strukturen er kjent som EF-hånden. EF-håndstrukturene er forbundet med hydrogenbindinger mellom de antiparallelle beta-arkene av calmodulin.
Funksjon, effekt og oppgaver
Calmodulin trenger tre til fire bundne kalsiumioner per molekyl for å være aktive. I den aktiverte tilstanden er det dannede kalsiumkalmodulinkomplekset involvert i reguleringen av et stort antall reseptorer, enzymer og ionekanaler med en rekke funksjoner. De regulerte enzymene inkluderer fosfatase-calcineurin, som spiller en viktig rolle i regulering av immunresponsen, og endotelial nitrogenoksydsyntase (eNOS), som produserer NO, som blant annet brukes til avslapning av glatte muskler og dermed for en utvidelse av Blodårer.
Ved lave kalsiumkonsentrasjoner aktiveres også adenylatcyklase (AC), ved høye kalsiumkonsentrasjoner, derimot, den enzymatiske motstykket, fosfodiesterase (PDE). På denne måten oppnås en kronologisk sekvens av reguleringsmekanismene: innledningsvis setter AC en signalbane i bevegelse via produksjonen av syklisk AMP (cAMP), senere slås dette av igjen av motstanderen PDE via cAMP-degradering. Imidlertid er den regulatoriske effekten av calmodulin på proteinkinaser som CaM-kinase II eller myosin lettkjede-kinase (MLCK), som vil bli forklart mer detaljert nedenfor, spesielt kjent.
CAMKII kan binde en fosfatrest til forskjellige proteiner og derved påvirke energimetabolismen, permeabiliteten for ioner og frigjøring av nevrotransmittere fra cellene. CAMKII finnes i spesielt høye konsentrasjoner i hjernen, der den spiller en viktig rolle i nevronal plastisitet, d.v.s. alle læringsprosesser. Men calmodulin er også uunnværlig for bevegelsesprosesser. I hvile er konsentrasjonen av kalsiumioner i en muskelcelle veldig lav og calmodulin er derfor inaktiv. Hvis muskelcellen er spent, strømmer kalsium imidlertid inn i celleplasmaet og okkuperer som en kofaktor de fire bindingsstedene på calmodulin.
Dette kan nå aktivere myosin lettkjedekinase, som forskyver de kontraktile fibrene i cellen og dermed muliggjør muskelsammentrekning. Andre, mindre kjente enzymer som er påvirket av calmodulin er guanylatcyklase, Ca-Mg-ATPase og fosfolipase A2.
Utdanning, forekomst, egenskaper og optimale verdier
Calmodulin forekommer i alle eukaryoter, som inkluderer alle planter, dyr, sopp og gruppen av amoeboid skapninger. Siden calmodulin-molekylet i disse organismer vanligvis er strukturert på en relativt lignende måte, kan det antas at det er et evolusjonært veldig gammelt protein som dukket opp på et tidlig tidspunkt.
Som regel er calmodulin til stede i relativt store mengder i plasma av en celle. I cytosol fra nerveceller er for eksempel den vanlige konsentrasjonen rundt 30-50 uM, d.v.s. 0,03-0,05 mol / L Proteinet blir dannet under transkripsjon og translasjon ved bruk av CALM-genet, hvorav det er tre alleler som er kjent til dags dato, som er referert til som CALM-1, CALM-2 og CALM-3.
Sykdommer og lidelser
Det er noen kjemikalier som kan ha en hemmende effekt på calmodulin og er derfor kjent som calmodulin-hemmere. I de fleste tilfeller er deres hemmende effekt basert på det faktum at de transporterer kalsium ut av cellen og dermed fjerner det fra calmodulin, som da bare er til stede i inaktiv tilstand.
Disse hemmende stoffer inkluderer for eksempel W-7. I tillegg hemmer noen psykotrope fenotiazinmedisiner calmodulin. Så vidt bredde som reguleringsfunksjonene til calmodulin, så forskjellige er de tenkelige feil og forstyrrelser når proteinet ikke lenger kan aktiveres av kofaktorkalsium og de regulerte målenzymer i seg selv er mindre aktive. Mangelfull aktivering av CAMKII, for eksempel, kan føre til en begrensning av den nevrale plastisiteten, som danner grunnlaget for læringsprosesser.
Minskende aktivering av MLCK svekker sammentrekningen av muskler, noe som kan føre til bevegelsesforstyrrelser. Mindre aktivering av enzymet calcineurin på grunn av en calmodulinmangel ville påvirke kroppens immunrespons og mindre aktivering av eNOene ville føre til lavere NO-konsentrasjoner. Sistnevnte forårsaker hovedsakelig problemer der nitrogenoksidet ellers er ment å forhindre uønsket blodkoagulering og utvide karene med det formål å bedre blodsirkulasjonen. Imidlertid bør det også nevnes på dette punktet at kalsiumsensoren Frequenin kan overta de biologiske funksjonene til calmodulin under visse forhold og dermed erstatte molekylet.






.jpg)