De eksitatorisk postsynaptisk potensiale er et spennende potensiale i den postsynaptiske membranen til nevroner. De individuelle potensialene oppsummeres romlig og midlertidig og kan dermed skape et handlingspotensial. Overføringsforstyrrelser som myastenia gravis eller andre myastenier forstyrrer disse prosessene.
Hva er det eksitatoriske postsynaptiske potensialet?

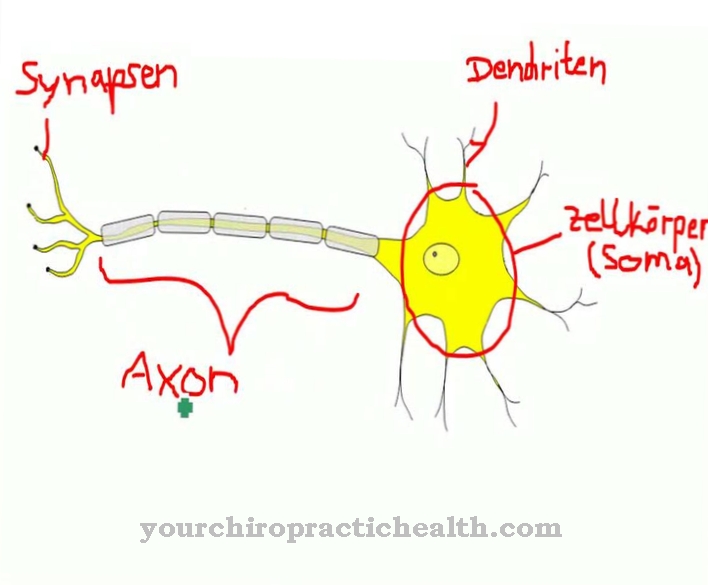
Nevroner skilles fra hverandre med et gap på 20 til 30 nm, også kjent som et synaptisk gap. Det er det minimale gapet mellom den presynaptiske membranregionen til en nevron og den postsynaptiske membranregionen i nedstrøms nervecelle.
Nevroner overfører eksitasjon. Derfor blir deres synaptiske gap overkoblet ved frigjøring av biokjemiske messenger-stoffer, som også er kjent som nevrotransmittere. Dette skaper et eksitatorisk postsynaptisk potensiale på membranområdet til nedstrømscellen. Det er en lokalt begrenset endring i det postsynaptiske membranpotensialet. Denne gradvise endringen i potensialet utløser et handlingspotensial i det postsynaptiske elementet. Det eksitatoriske postsynaptiske potensialet er en del av den neuronale eksitasjonsledningen og oppstår når nedstrøms cellemembran er depolarisert.
De spennende postsynaptiske potensialene blir mottatt og behandlet av følgende nevron ved å legge opp både romlig og midlertidig. Når cellens terskelpotensiale overskrides, føres et nydannet handlingspotensial bort av aksonet.
Det motsatte av det eksitatoriske postsynaptiske potensialet er det hemmende postsynaptiske potensialet. Dette fører til hyperpolarisering på den postsynaptiske membranen, som forhindrer utløsing av et handlingspotensial.
Funksjon & oppgave
Det spennende postsynaptiske potensialet og det hemmende postsynaptiske potensialet påvirker alle nerveceller. Når terskelpotensialet deres overskrides, depolariseres nerveceller. De reagerer på denne depolarisasjonen ved å frigjøre eksitatoriske nevrotransmittere. En viss mengde av disse stoffene aktiverer de sendersensitive ionekanalene i nevronen. Disse kanalene er permeabel for kalium- og natriumioner. Lokale og graderte potensialer i betydningen et eksitatorisk potensial depolariserer dermed den postsynaptiske membranen til nevronen.
Når membranpotensialet avledes intracellulært, er det eksitatoriske postsynaptiske potensialet depolarisering av soma-membranen. Denne depolarisasjonen skjer som et resultat av passiv forplantning. Det er en summering av individuelle potensialer. Mengden av frigitt nevrotransmitter og størrelsen på det rådende membranpotensialet bestemmer omfanget av det eksitatoriske postsynaptiske potensialet. Jo høyere pre-depolarisering av membranen, jo lavere er det eksitatoriske postsynaptiske potensialet.
Hvis membranen allerede er depolarisert over sitt hvilepotensial, faller det postsynaptiske eksitasjonspotensialet og når det under visse omstendigheter null. I dette tilfellet oppnås reverseringspotensialet for det eksiterende potensialet. Hvis pre-depolarisasjonen viser seg å være enda høyere, oppstår et potensiale med et motsatt tegn. Dermed må ikke det eksitatoriske postsynaptiske potensialet likestilles med en depolarisering. Den beveger membranen heller mot et visst likevektspotensial, som ofte forblir under det respektive hvilemembranpotensialet.
Arbeidet med en kompleks ionemekanisme spiller en rolle i dette. Med det eksitatoriske postsynaptiske potensialet kan en økt membranpermeabilitet for kalium- og natriumioner observeres. På den annen side kan potensialer med redusert konduktivitet for natrium- og kaliumioner også oppstå. I denne sammenhengen antas ionekanalmekanismen å være utløsningen for lukking av alle lekk kaliumionkanaler.
Det hemmende postsynaptiske potensialet er det motsatte av det eksitatoriske postsynaptiske potensialet. Også her endrer membranpotensialet lokalt på den postsynaptiske membranen til nerveceller. Hyperpolarisering av cellemembranen skjer ved synapsen, som hemmer utløsningen av handlingspotensialer innenfor rammen av det eksitatoriske postsynaptiske potensialet. Nevrotransmitterne ved de hemmende synapser utløser en cellerespons. Kanalene i den postsynaptiske membranen åpner og lar kalium- eller kloridioner passere. Den resulterende innstrømningen av kaliumion og kloridion forårsaker lokal hyperpolarisering i den postsynaptiske membranen.
Du finner medisinene dine her
➔ Medisiner for muskelsvakhetSykdommer og plager
Ulike sykdommer forstyrrer kommunikasjonen mellom individuelle synapser og dermed også signaloverføringen ved den kjemiske synapsen. Et eksempel er den nevromuskulære sykdommen myasthenia gravis, som påvirker muskelendeplaten. Det er en autoimmun sykdom av tidligere ukjent årsak. Ved sykdommen danner kroppen autoantistoffer mot kroppens eget vev. Ved muskelsykdom er disse antistoffene rettet mot den postsynaptiske membranen på nevromuskulære endeplater. Oftest er autoantistoffene i denne sykdommen acetylkolinreseptorantistoffer. De angriper nikotinacetylkolinreseptorene på forbindelsespunktene mellom nerver og muskler. Den resulterende immunologiske betennelsen ødelegger det lokale vevet.
Som et resultat forstyrres kommunikasjonen mellom nerve og muskel, siden interaksjonen mellom acetylkolin og dens reseptor blir vanskeliggjort eller til og med forhindret av acetylkolinreseptorantistoffene. Handlingspotensialet kan derfor ikke lenger passere fra nerven til muskelen. Muskelen er derfor ikke lenger spennende.
Summen av alle acetylkolinreseptorer reduseres samtidig som reseptorene blir ødelagt av immunaktiviteten. De subsynaptiske membranene går i oppløsning og endocytose skaper et autofagosom. Transportvesikler smelter sammen med autofagsomene og acetylkolinreseptorene endres som et resultat av denne immunreaksjonen. Med disse endringene endres hele motorens endeplate. Det synaptiske gapet utvides. Av denne grunn diffunderer acetylkolin fra synaptisk spalte eller hydrolyseres uten å binde seg til reseptoren.
Andre myastenier viser lignende effekter på synaptisk spalte og det eksitatoriske postsynaptiske potensialet.









.jpg)
.jpg)
.jpg)
.jpg)